Overview
I made a twitter post today which read: "Evolutionary theory does NOT indicate a human male & female pair that is a bottleneck, common ancestor: True or False?"
That was the best I could do in 140 characters but I think I captured the essence of the question. The main purpose of this was to challenge people to think about how such evolutionary processes might work.
To explain in a bit more detail - the question is, for speciation to occur does a M/F pair of the NEW species need to pre-exist or not. If we cannot breach this barrier then evolution would be dead in the water.
I also have found this is an extremely common misconception amongst people who do not understand evolution that there is this requirement for a new species to be 'magically' created in pairs. They don't seem to understand that speciation happens in a population of viable procreation partners.
And while it is possible, in theory, that two individuals could just happen to have the SAME genetic mutation at the same time AND find each other it is simply not necessary for evolutionary speciation to occur (and co-mutation seems unlikely to be the norm).
So, the correct answer is True - evolutionary theory does NOT predict or require that there would be M/F pairs created for speciation to occur. Speciation is really the product of culling the population rather than procreation. That is, the isolated species is created when other 'bridge' members of the population die, rather than when it is born.
Thought Experiment:
If you isolated 2 groups of 100 rabbits (call them A and B) each then wait 200My with each group exposed to different selectivity criteria (e.g., group A in harsher, colder climates and group B in warmer) what would we expect to find? Well, the A' rabbits would all be fairly similar to each other but very different from the A rabbits - they might have smaller ears, warmer fur, cold-adapted respiratory systems, etc. Likewise, the B' rabbits would be similar to each other but again different from the original group B. They might have lost their dense fur for example.
And let's say that you saved some embryo's from the original population and raise them into 'normal' rabbits - these original rabbits might be completely unable to bred with either A' or B' (each generation spent in isolation would slightly increase this chance).
Each group will likely have speciated over that time but at no point would there be a unique M/F PAIR in the diverged lineages. They would ALL be related to each other with finer gradations of genetic variations.
Detailed Example
This next one is very technical but will demonstrate, I hope very clearly, what happens in a population leading up to speciation and why a M/F pair isn't required for the process.
First, please note that this account is FICTIONAL, although it is based on science I'm essentially paraphrasing. For example, there really IS an OCX family of genes which make Ovocalyxin proteins which are involved in egg-shell production (search for OCX-32, OCX-36, OCX-21 if you want to learn more). But I'm calling OCX1 my baseline and just numbering the successive mutations, I'm not using the SAME numbering scientists use.
We will look at three gene families (groups of genes) in a population of Gray junglefowl (the probable ancestor of the domesticated chicken) and we will crack the old chicken and egg problem (please hold your groans until the end).
Gene #1 will be OCX in the Ovocalyxin family of genes and is related to egg-shell production
Gene #2 will be SLIT, an axon guidance molecule in embryonic development (again, a real gene designation - but fictional mutations for purpose of demonstration)
Gene #3 will be MHC major histocompatibility complex - which controls the antigens affecting immune response; there are many possible sources of evolved sexual incompatibility; I'm using an antigen example
In each case, there exists a fairly large, and viable set of variants for these genes. We will number the variants start with 1 as the baseline for our population of Gray junglefowls as {1,1,1} which means OCX1, SLIT1, MHC1 while {1,1,2} has mutation variant MHC2.
In our make-believe world, the Gray junglefowls have softer-shelled eggs which are easily poached (no groans!) by snakes and other small animals, the eggs take 28 days to mature and hatch, and the animals have compatible antigen systems.
Also, imagine in our population there are 1000's of fowl although we will focus on just a few individuals. I will call each fowl F and a number (e.g., F1, F2, F3).
Our story begins when F1{1,1,1} mates with F2{1,1,1} and has offspring F3{1,2,1}. This lucky fowl develops and hatches in only 26 days, giving less time for predation to occur resulting in a slightly increased chance of survival.
F3{1,2,1} grows up and mates with F4{1,1,1} giving several Fn{1,1,1} offspring and two fowl F5{1,2,1} and F6{1,2,1}. The Fn's are eaten before they hatch but F5 & F6 survive due to the shorter 26 day incubation period thanks to the mutation to their SLIT gene complex. Hopefully it is easy to see how this trait could easily spread through the population. Year after Year, SLIGHTLY more of the {1,2,1} variants survive until finally the distribution is 95%{1,2,1} and only 5%{1,1,1}.
Is this minor mutation a new species? Well no, they can all freely interbred and while we think of Gray1 as having a 28 day incubation period and the 26 day incubation period of Gray2 alone doesn't seem like enough to call it a new species. Thanks only to the power of genetic sequencing we can SEE the exact change that occurred between the two groups in the population.
Possibly 100's of more years go by -- other genes mutate, maybe they get bigger or smaller feathers or their colors shift slightly - but still nothing we consider 'major' and still all sexually compatible with each other.
Finally, a single F7{2,2,1} is born and happens to survive. When F7 lays an egg is has a tougher shell than eggs from other Gray's! This harder shell thwarts several of the predators of the softer shells. The {2,2,1} mutation spreads quickly through the population as not only do these fowl survive predation it forces the predators to eat more of the {1,2,1} and {1,1,1} fowl.
The population quickly (in geological time) changes to 95%{2,2,1}, 3%{2,1,1}, 1.5%{1,2,1}, 0.5%{1,1,1} - almost all of the SLIT1 variants are now gone having been eaten before hatching either by taking too long or being easier targets.
There are likely MANY genetic variations which prove unviable, these fowl die before they are born or are unable to reproduce. Time marches forward. It is here where we need to understand that fowls are Diploid genetically (they have two copies) and thus can express dominate or recessive traits. I will mark genes which are different on the chromosome with a prime (e.g., 1'2 means it has both 1 and 2 variants).
Our next shift involves the MHC complex - this is the true beginning of our speciation event. F8{2,2,2'} is born, this minor shift in the MHC complex gives this variant a slightly lower chance of successful sexual reproduction (but possibly an increased resistance to a common disease).
F8{2,2,1'2} can then mate with F7{2,2,1}s giving both types of offspring (those like F8 and those like F7). At first, there would be no mating issue as MOST mates would be compatible, so the variant is not detrimental at this stage and it is over the long haul where genetic variations compete with one another. Even seemingly detrimental variants which do not prohibit reproduction can flourish for a time.
Eventually our fowl friend will meet another like itself and one F8{2,2,1'2} will mate with another giving three possible offspring:
F7{2,2,1} 25%
F8{2,2,1'2} 50% (1'2 and 2'1 are the same)
F9{2,2,2} 25%
And here is where our new variant encounters a slight challenge. F9's cannot mate successfully with F7 or lower (their MHC complexes are too incompatible due to the now dominate MHC2) and only half the time with F8's.
But we can see from the above that F8's are favored to become the most frequent so our fine feathered fowl has a decent chance of making it. And, unlike the mutation event itself, we do NOT have only one shot here. There can fairly quickly be 100's or thousands of F9's trying to mate and their chances are better than it might seem (50% chance with 50% of the population and 100% chance with 25%). And if there is some ancillary benefit to {2,2,2} like disease resistance that could kick it over the top.
Let's even say it has failed 1000 times before but THIS time it makes it. We suddenly have a very robust population of fowl with very diverse genomes but with a peak of 90%{2,2,2} and 8%{2,2,1'2} fowl.
So now you ask, where are the Chickens? These are all just Gray's right? Well, it is a Gray with a hardened shell, faster incubation, and sexual incompatibility with any {1,1,1} population that was perhaps isolated from our earlier group.
Let's say there are further enhancements {3,2,2} and {2,3,2} which result in a higher population of {3,3,2}'s and eventually {4,4,2}'s, {4,4,2'3}s and {4,4,3}s which are even more disease resistant. All these changes using the same processes above.
So now we have a fowl {4,4,3} with an extremely tough & bacteria resistant shell, a 21-day incubation period, and is more robust against diseases. This FIRST {4,4,3} is our first True Chicken. However, it is still sexually compatible, at various levels with the {4,4,2'3} fowl which are not quite True Chickens.
We now require only ONE more piece of information and we can resolve the famous Chicken vs Egg Dilemma. What do you mean by 'egg'?
a) An egg that CONTAINS a True {4,4,3} Chicken - the egg which contained the first True Chicken would have come from two {4,4,2'3}-Almost Chickens -- therefore the Egg would have come first.
b) An egg FROM a True {4,4,3} Chicken - then the True {4,4,3} Chicken must have come first
c) Any type of egg at all - eggs in general are much much older than chickens of all kinds
Supporting Data
If you chart out the allele frequencies over tme you get a graph that looks like this (from Wiki):
{kind=link}
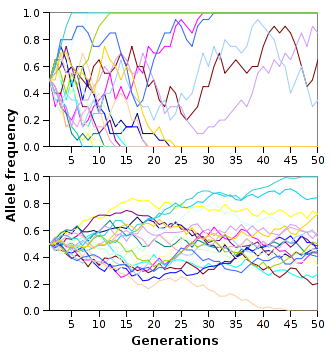
Here is a very nice study on allele frequency distribution in geographically diverse populations of mice.
Another study in which e coli, under observed laboratory conditions, evolved the ability to process citrate with very detailed genomics data through the generations.
Learn about the HOX gene complex, which is a major factor in developmental morphology (and some cool examples in Drosophila).
Meet Prdm9 (PR domain-containing 9), the gene responsible for much of the genetic shuffling that is done when sperm eggs develop from germ cells (requires NewScientist account). This gene comes in many variations which alter how much shuffling is done and at which points (and some variations may be involved in certain kinds of genetic diseases) - more on Prdm9. Nature Genetics journal paper: doi:10.1038/ng.658
Learn about Histone and its role in shaping and regulating DNA expression.
A critical step in the fertilization process is Oocyte activation, within this process is the DNA synthesis from the two gametes (called syngamy, this combines the DNA from each gamete) and there are complexes of genes which must interlock or the synthesis will not happen, even if the DNA is otherwise compatible. This is one of the mechanisms of speciation by sexual incompatibility in complex organisms (penis shape and size is another in some species, e.g., ducks). Differences in these genes are involved in some cases of human infertility which could eventually lead to speciation in humans.
Interesting paper on the speciation of Yeast due to reproductive isolation:
Incompatibility of Nuclear and Mitochondrial Genomes Causes Hybrid Sterility between Two Yeast Species.
No comments:
Post a Comment